13 Neurotransmitter Action: Ionotropic Receptors
Ionotropic receptors, also called neurotransmitter-gated or ligand-gated channels, are ion channels that open in response to the binding of a neurotransmitter. They are primarily located along the dendrites or cell body, but they can be present anywhere along the neuron if there is a synapse. Ligand-gated channels are important for receiving incoming information from other neurons.
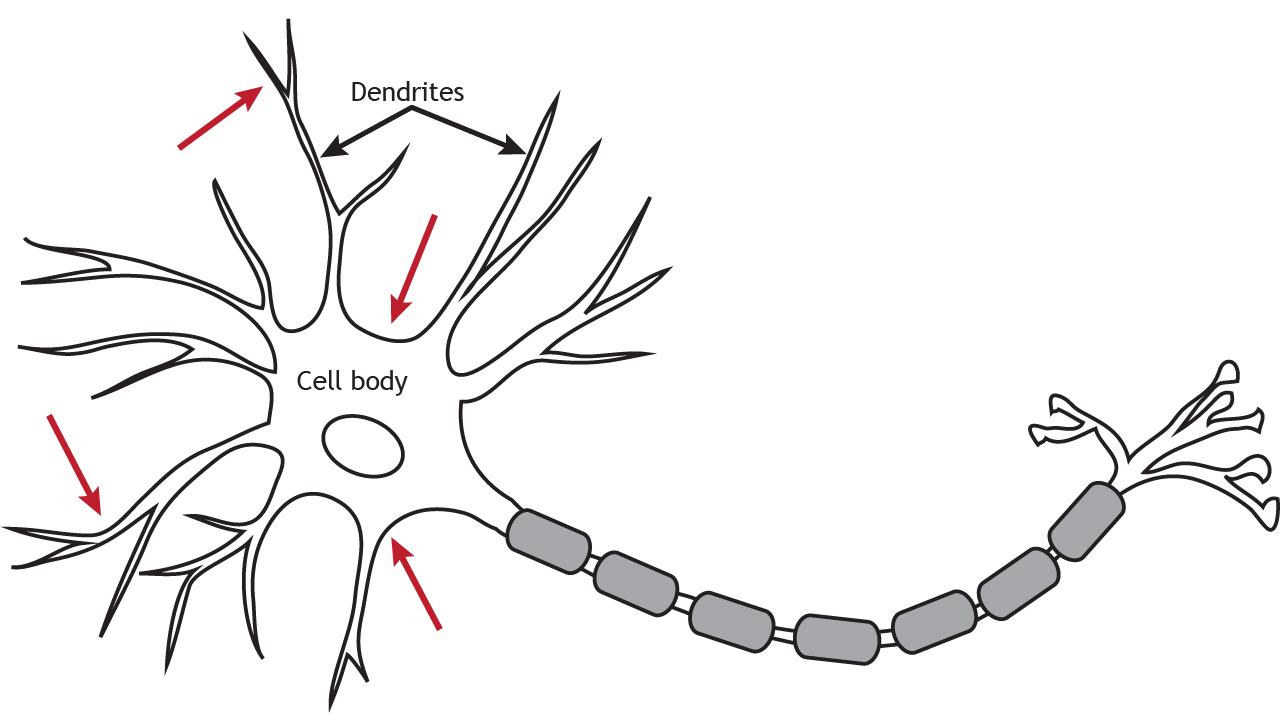
Although ionotropic receptors are ion channels, they open in a different way than the voltage-gated ion channels needed for propagation of the action potential. The ionotropic receptors are ligand-gated, which means that a specific molecule, such as a neurotransmitter, must bind to the receptor to cause the channel to open and allow ion flow. As seen in previous chapters, the voltage-gated channels open in response to the membrane potential reaching threshold.
Animation 13.1. Ionotropic receptors, also called ligand-gated channels, are ion channels that are opened by the binding of neurotransmitters. Voltage-gated channels are opened by the membrane potential of the cell reaching threshold. Both types of channels allow ions to diffuse down their electrochemical gradient. The lined, teal channels represent glutamate receptors; the solid yellow channels represent GABA receptors; the dotted, blue channels represent voltage-gated sodium channels. ‘Ion Channel Gating’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
The receptors can only be opened by a specific ligand. Neurotransmitters and receptors fit together like a lock and key; only certain neurotransmitters are able to bind to and open certain receptors.
Animation 13.2. Since neurotransmitter receptors can only bind specific neurotransmitters, glutamate binds to and opens glutamate receptors but has no effect on GABA receptors. The lined, teal channels represent glutamate receptors; the solid yellow channels represent GABA receptors. ‘Ligand and Receptor’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Ion movement through ligand-gated ion channels follows the same principles covered in previous chapters
Glutamate Receptors
Glutamate causes EPSPs by opening cation channels that increase sodium permeability across the membrane
Glutamate is the primary excitatory neurotransmitter in the central nervous system and opens non-selective cation channels. There are three subtypes of glutamate receptors. The AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) and kainate receptors allow both sodium and potassium to cross the membrane. Although potassium can leave the cell when the receptors open, the electrochemical gradient driving sodium ion movement is stronger than the gradient driving potassium movement, resulting in a depolarization of the membrane potential.
Animation 13.3. AMPA and kainate glutamate receptors are non-selective ion channels that allow both sodium and potassium to flow across the membrane. When glutamate binds, sodium flows in and potassium flows out. The lined, teal channel represent sAMPA receptors; the checkered, teal channel represents kainate receptors. ‘AMPA and Kainate’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
The NMDA (N-methyl-D-aspartate) receptor requires the binding of glutamate to open, but it is also dependent on voltage. When the membrane potential is below, at, or near rest, a magnesium ion blocks the open NMDA receptor and prevents other ions from moving through the channel. Once the cell depolarizes, the magnesium block is expelled from the receptor, which allows sodium, potassium, and calcium to cross the membrane. The voltage change needed to open the NMDA receptor is usually a result of AMPA receptor activation. Released glutamate binds to both AMPA and NMDA receptors, sodium influx occurs through open AMPA channels, which depolarizes the cell enough to expel the magnesium ion and allow ion flow through the NMDA receptors.
Animation 13.4. NMDA receptors are opened by a combination of glutamate binding and a voltage trigger. At low levels of stimulation, when the the membrane potential is near rest, a magnesium ion blocks the open NMDA receptor channel preventing ion flow. Ions can flow through open AMPA receptors, which begins to depolarize the membrane. The voltage change eventually expels the magnesium ion from the channel, allowing sodium, potassium, and calcium to cross the membrane. The lined, teal channel represents AMPA receptors; the dotted, violet channel represents NMDA receptors. ‘AMPA and NMDA’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Nicotinic Acetylcholine Receptors
Like glutamate receptors, nicotinic acetylcholine receptors are non-selective cation channels. Nicotinic receptors, though, are located primarily outside of the central nervous system and are primarily used at the neuromuscular junction.
GABA and Glycine Receptors
GABA and Glycine cause IPSPs by opening chloride channels that increase chloride permeability across the membrane
GABA and glycine receptors are chloride channels. Since an increase chloride permeability across the membrane is inhibitory, the binding of GABA or glycine to their respective ionotropic receptor will cause inhibition.
Animation 13.5. GABA and glycine are inhibitory receptors that are selective to chloride. The solid yellow channel represents a GABA receptor; the patterned, yellow channel represents a glycine receptor. ‘GABA and Glycine’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Ionotropic Receptors Cause Postsynaptic Potentials
Postsynaptic potentials (Chapter 5) are a result of ionotropic receptors opening. Excitatory ionotropic receptors increase sodium permeability across the membrane, whereas inhibitory ionotropic receptors increase chloride permeability. Ion flow through the ionotropic receptors follows the same principles as other ion channels covered so far.
Equilibrium Potential Review
Previously, we covered ion movement through voltage-gated channels and discussed that electrochemical gradients will drive ion movement toward equilibrium. The neuron’s membrane potential at which the chemical and electrical gradients balance and equilibrium occurs is the ion’s equilibrium potential.
Animation 13.6. Ions move through open voltage-gated channels trying to reach equilibrium. As the ions cross the membrane, the neuron’s membrane potential moves closer to the ion’s equilibrium potential. In the animation, a voltage-gated sodium channel opens, and sodium flows in until the membrane potential equals approximately +60 mV, sodium’s equilibrium potential. The blue, dotted channel represents a voltage-gated sodium channel. ‘Equilibrium Potential’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Reversal Potential
This same principle is used for ion movement through ionotropic receptors. The membrane potential at which ion flow through a receptor is at equilibrium is called the reversal potential of the receptor. The direction of ion movement can be predicted if the reversal potential of the receptor is known.
You can think of a reversal potential as being the equilibrium potential for a receptor, which may allow more than one ion to move across the membrane
GABA and Glycine – Receptors Selective to One Ion
When an ionotropic receptor that is selective to only one ion opens, the reversal potential of the receptor is the same as the equilibrium potential of the ion. GABA and glycine receptors only allow chloride ions to cross the membrane. Therefore, the reversal potential of a GABA or glycine receptor is equal to the equilibrium potential of chloride, and the binding of GABA or glycine to their respective ionotropic receptor will cause an inhibitory postsynaptic potential (IPSP).
Animation 13.7. Ions move through open ligand-gated channels trying to reach equilibrium. As the ions cross the membrane, the neuron’s membrane potential moves closer to the receptor’s reversal potential. When the ionotropic receptor only increases permeability for one ion, the receptor’s reversal potential is the same as the ion’s equilibrium potential. In the animation, a GABA receptor open, and chloride flows in until the membrane potential equals approximately -65 mV, GABA’s reversal potential and chloride’s equilibrium potential. Increased chloride permeability causes an IPSP and inhibits the neuron. The yellow, checkered channel represents a GABA receptor. ‘GABA Reversal Potential’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Glutamate – Reversal Potential for Receptors that are Non-Selective
However, if the ionotropic receptor allows the flow of more than one ion, or is non-selective, the reversal potential of the receptor does not equal the equilibrium potential of either ion but is somewhere in between. The equilibrium potential of sodium is approximately +60 mV, and the equilibrium potential of potassium is approximately -80 mV. A glutamate receptor is a non-selective cation channel that allows the flow of both ions, and the reversal potential of the receptor is 0 mV. This means that if the neuron’s membrane potential is negative, the driving forces acting on sodium are stronger than the driving forces acting on potassium, so more sodium will flow in than potassium will flow out, and the membrane potential will depolarize, causing an excitatory postsynaptic potential (EPSP).
Animation 13.8. The reversal potential of an ionotropic receptor that is not selective to one ion will fall between the equilibrium potentials of the permeable ions. Glutamate receptors allow the flow of both sodium and potassium ions, so the reversal potential for the receptor is approximately 0 mV. More sodium will flow into the cell than potassium flows out, resulting in a depolarization of the membrane. The line, teal channel represents a glutamate receptor. ‘Glutamate Reversal Potential – Rest’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
If the membrane potential reached the reversal potential of the glutamate receptor, the electrochemical gradients acting on sodium and potassium would balance, so overall ion flow in both directions would be equal, and the membrane potential would not change.
Animation 13.9. At the reversal potential, there is no net ion flow in either direction. An equal number of sodium ions enter the cell as potassium ions leave. Since there is no change in voltage at the reversal potential, if the receptor remained open, the membrane potential would stay at 0 mV. ‘Glutamate Reversal Potential – 0 mV’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Key Takeaways
- Ionotropic receptors are ligand-gated ion channels that open when a specific neurotransmitter binds
- For receptors selective to one ion, the reversal potential equals the ion’s equilibrium potential
- For receptors not selective for only one ion, the reversal potential is a value between the ions’ equilibrium potentials
- Glutamate is an excitatory neurotransmitter that opens non-selective cation channels that allow the influx of sodium, causing an EPSP
- GABA and glycine are inhibitory neurotransmitters that open chloride channels, causing an IPSP
Test Yourself!
The following questions refer to a mythical cell, the Thinking Cell. The properties for this cell are given in the table. In all cases, the postsynaptic membrane is a dendrite. Use the information in the table to answer the following questions.
| Ion | Inside concentration (mM) | Outside concentration (mM) | Equilibrium Potential (mV) |
|---|---|---|---|
| A- | 6 | 125 | -65 |
| B+ | 12 | 120 | +60 |
| D+ | 125 | 5 | -84 |
| E++ | 0.00001 | 1.5 | +155 |
Video Lecture
Attributions
This chapter was adapted from “Neurotransmitter Action: Ionotropic Receptors” in Foundations of Neuroscience by Casey Henley which is licensed under a Creative Commons Attribution NonCommerical ShareAlike 4.0 International License.
